
El Dr. Dunn es investigador en el Centro de Ciencias Genómicas de la Universidad Nacional Autónoma de México, en Cuernavaca, Morelos. Estudia la fisiología bacteriana, con un enfoque en la interacción entre bacterias y plantas. Es miembro de la Academia de Ciencias de Morelos.
Esta publicación fue revisada por el comité editorial de la Academia de Ciencias de Morelos.
La primera parte de este ensayo [2] explicó cómo una fusión física, o endosimbiosis, entre microbios primitivos posibilitó la evolución de células complejas (eucariotas) que contienen fábricas productoras de energía llamadas mitocondrias. Esta endosimbiosis ocurrió hace más de dos mil millones de años y posibilitó la evolución de animales, plantas, hongos y protistas (eucariotes unicelulares) a partir de un ancestro eucariote común. La aparición de las mitocondrias, junto con la oxigenación de la atmósfera terrestre, aceleró considerablemente el ritmo y el alcance de la evolución. Aquí, en la segunda parte del ensayo, exploraremos cómo nuestro concepto de endosimbiosis como un motor de la evolución fue revitalizado hace casi 60 años por una bióloga inconformista llamada Lynn Margulis (Figura 1) y cómo nuestra comprensión de este proceso y su impacto en la diversidad de la vida en la Tierra se ha expandido desde entonces.
Cómo una antigua endosimbiosis dio origen a las mitocondrias
En su ambicioso artículo de 1967, "Sobre el Origen de las Células en Mitosis" ("On the Origin of Mitosing Cells"), la bióloga Lynn Margulis (escribiendo como Lynn Sagan) revivió la casi olvidada teoría de la endosimbiosis como un mecanismo evolutivo. El término "células en mitosis " en el título es otra forma de referirse a las células eucariotas. En su extenso trabajo, Margulis entrelazó evidencia moderna e histórica obtenida de la biología celular, la bioquímica, la paleontología y las ciencias de la Tierra para sustentar su afirmación de que "...la célula eucariota es el resultado de la evolución de antiguas simbiosis" [12]. Argumentó que las mitocondrias en las células de todos los eucariotes y los cloroplastos en los eucariotes fotosintéticos surgieron de eventos endosimbióticos separados.
Las ideas sobre la endosimbiosis como progenitora de los organelos eucariotes tienen una larga historia. A finales del siglo XIX, el patólogo alemán Richard Altmann (Figura 1) creía que las mitocondrias que observaba dentro de las células eran organismos vivos autónomos, a los que llamó bioblastos. En su artículo, Margulis reconoció a los biólogos Constantin Mereschkowski, de Rusia, e Ivan Wallin, de Estados Unidos (Figura 1), por sus propuestas sobre la endosimbiosis como posible origen de ciertos organelos en las células eucariotas [12]. Mereschkowski propuso en 1905 que los cloroplastos eran "cianobacterias reducidas" y que la capacidad fotosintética de las plantas fue heredada completamente de las cianobacterias. Wallin, trabajando en la década de 1920, planteó la hipótesis de que las mitocondrias habían sido bacterias de vida libre, basándose en similitudes en su apariencia y composición química. En las décadas posteriores al trabajo en gran medida ignorado de estos científicos, se descubrió que las mitocondrias de las células eucariotas tenían algunas características en común con las bacterias, incluyendo su tamaño, forma y su modo de reproducción por fisión binaria (cuando una célula se divide en dos al partirse por la mitad). Sin embargo, los métodos científicos de aquella época no eran capaces de proporcionar evidencia sólida para sus afirmaciones, que fueron en gran medida olvidadas hasta que Margulis las revitalizó y amplió.

Figura 1. Algunos de los científicos que aportaron conceptos clave a la teoría endosimbiótica.
Para explicar el origen de las mitocondrias, Margulis propuso que una bacteria con metabolismo aeróbico, capaz de usar oxígeno para producir energía de manera eficiente, fue engullida por una bacteria anaerobia, cuya incapacidad para usar oxígeno la hacía energéticamente deficiente [12]. Recordemos que hace más de dos mil millones de años, las cianobacterias fotosintéticas comenzaron a oxigenar la atmósfera de la Tierra (el Gran Evento de Oxigenación o GOE), mucho antes de la llegada de los eucariotes [8]. La bacteria aeróbica se estableció como un endosimbionte dentro de la célula hospedadora anaerobia, dándole un metabolismo más eficiente y una ventaja competitiva frente a otras bacterias anaeróbicas. El metabolismo aeróbico no sólo proporcionó a las células un gran impulso energético, sino que su consecuencia natural fue que el oxígeno de las células se consumió, lo que es importante porque el oxígeno es bastante tóxico para muchas organismos anaeróbicos. Con el tiempo, los sistemas genéticos y metabólicos del endosimbionte se integraron tanto con los del hospedador, que el endosimbionte se transformó gradualmente en una mitocondria (Figura 2A.) [4].
Todas las células obtienen su energía al descomponer nutrientes mediante una serie de reacciones químicas, cuyo resultado final es la formación de ATP (trifosfato de adenosina), la principal moneda energética de la célula. Cuando la célula necesita energía, el ATP se transforma y libera la energía necesaria para funciones como el metabolismo, el transporte de sustancias o el movimiento celular.
Como se explica con más detalle en la Figura 2B, la respiración aeróbica en las células eucariotas comienza con la glucosa, una molécula rica en energía química que se metaboliza para generar moléculas (NADH y FADH₂) que contienen electrones ricos en energía. En la mitocondria, la energía de estos electrones se utiliza para mover iones de hidrógeno (protones o H+) de un lado a otro de la membrana mitocondrial interna. Los protones fluyen desde una región de alta concentración en un lado de la membrana hacia una región de menor concentración en el otro. Para ello, pasan a través de la ATP sintasa, una enzima unida a la membrana que actúa como la rueda hidráulica en una presa. Este flujo de protones impulsa la ATP sintasa para producir ATP (Figura 2B).
Las mitocondrias producen la mayor parte del ATP en las células eucariotas, que pueden tener cientos o miles de mitocondrias por célula, todas ellas realizando respiración aeróbica. Las membranas internas de las mitocondrias siguen una ruta muy intrincada, lo que les da una gran superficie (Figura 2A). Como resultado, cada mitocondria tiene una gran cantidad de espacio cubierto por proteínas de la cadena de transporte de electrones y ATP sintasa (Figura 2B). El resultado es que las mitocondrias pueden producir mucho ATP.
Las mitocondrias realizan respiración aeróbica mediante un sistema que se originó en las bacterias, mucho antes de que existieran las mitocondrias. Sin embargo, a diferencia de las mitocondrias, la membrana interna de las bacterias que alberga su sistema de transporte de electrones y la ATP sintasa no está tan firmemente plegada y, por lo tanto, contiene muchas menos de estas proteínas. Esto significa que las células procariotas son comparativamente deficientes en energía en comparación con las células eucariotas, aunque las estimaciones sobre la magnitud de esta diferencia varían ampliamente [5].
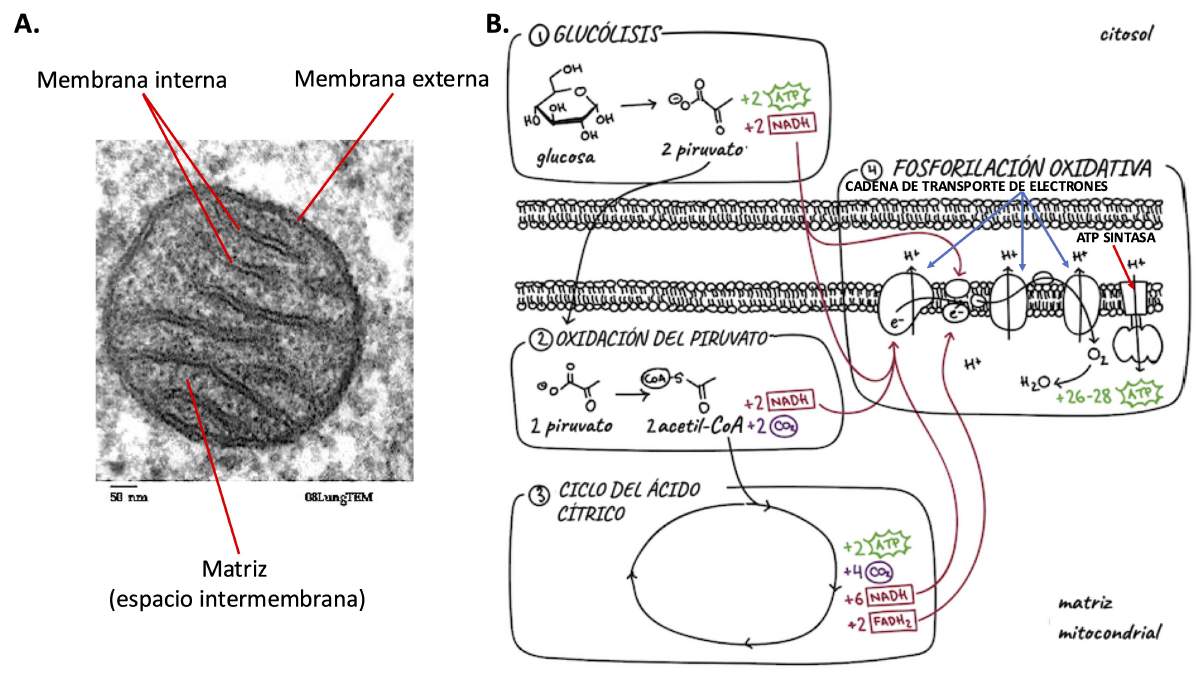
Figura 2. A, Mitocondria de una célula de mamífero. B, Cómo se produce el ATP por fosforilación oxidativa en las mitocondrias. Paso 1. La vía de la glucólisis convierte la glucosa en piruvato en el citosol de la célula. Luego, el piruvato es transportado al interior de la matriz mitocondrial. Paso 2. En la matriz mitocondrial, el piruvato se transforma en acetil-CoA, la molécula que inicia el ciclo del ácido cítrico. Paso 3. El acetil-CoA es metabolizado por el ciclo del ácido cítrico, lo que produce los transportadores de electrones FADH₂ y NADH. Paso 4. Las reacciones en las que estos transportadores donan electrones a la cadena de transporte de electrones también generan protones (H⁺). Estos protones son bombeados fuera de la matriz hacia el espacio entre la membrana mitocondrial interna y la membrana externa, donde se acumulan. Para regresar a la matriz (movilizándose a favor del gradiente de concentración, desde el lado con alta concentración de H⁺ hacia el lado con baja concentración de H⁺), los protones deben pasar a través de la ATP sintasa, lo que impulsa la producción de ATP. Parte A modificada de https://en.wikipedia.org/wiki/File:Mitochondria,_mammalian_lung_-_TEM.jpg. Parte B es bajo una licencia de Creative Commons 4.0 Internacional.
A partir que las células eucariotas tienen volúmenes muchas veces mayores que las células procariotas, la cantidad de ATP producida por sus numerosas mitocondrias les proporciona la energía suficiente para alimentar procesos internos de transporte, movimiento, reproducción y el mantenimiento de sus grandes genomas. Su gran tamaño y la capacidad de engullir organismos más pequeños, como bacterias, permitieron que algunos eucariotes se convirtieran en depredadores efectivos [5].
Una segunda endosimbiosis que transformó la vida en la Tierra ocurrió unos 600 millones de años después de la evolución de las mitocondrias. Margulis planteó la hipótesis que una célula eucariota engulló a una cianobacteria fotosintética, que se estableció y evolucionó hasta convertirse en un cloroplasto [10,12] (Figura 3). De esta endosimbiosis surgieron las algas y las plantas terrestres verdes. Gracias a sus cloroplastos, las plantas y las algas pueden generar energía a partir de la luz solar, el agua y el dióxido de carbono, y son, por tanto, productores primarios vitales en la biosfera.
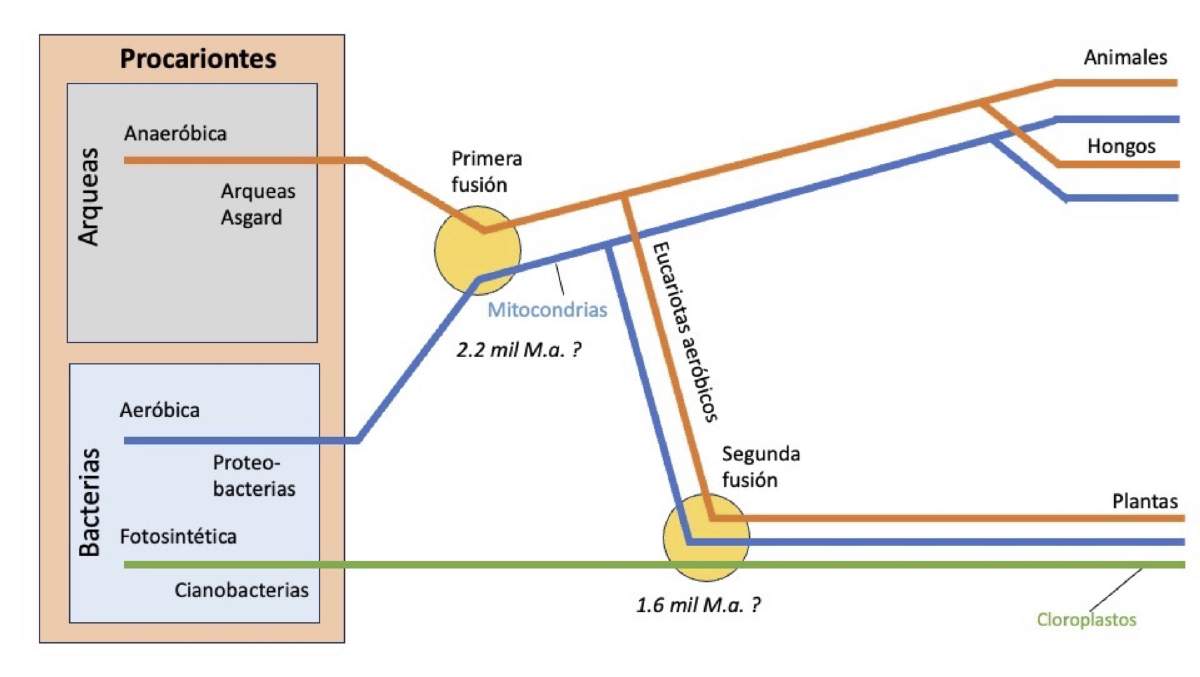
Figura 3. Línea del tiempo de los eventos endosimbióticos (o fusiones) que dieron origen a las mitocondrias y los cloroplastos. La primera fusión fue entre un endosimbionte bacteriano aeróbico y un hospedero arqueano anaeróbico. Con el tiempo, el endosimbionte evolucionó hasta convertirse en una mitocondria. Las líneas azul y naranja que emergen de esta primera fusión representan linajes con características genéticas tanto de proteobacterias (mitocondrias) como de arqueas, respectivamente. Por eso, animales y hongos son organismos quiméricos, con genes provenientes de ambos dominios. La segunda fusión ocurrió entre un eucariote aeróbico y una cianobacteria (línea verde), y esta última terminó evolucionando en cloroplastos fotosintéticos. Las plantas también son organismos quiméricos, con aportes genéticos de linajes proteobacterianos, cianobacterianos y arqueanos. Redibujado y traducido a partir de la figura mostrada en: https://en.wikipedia.org/wiki/Symbiogenesis.
Desde Margulis: Nuevas ideas sobre el origen de las mitocondrias y cloroplastos
Inicialmente, la formulación de Margulis sobre el origen endosimbiótico de las mitocondrias y los cloroplastos enfrentó una gran resistencia, pero fue reivindicada en las décadas siguientes a medida que se acumulaba una abrumadora evidencia en apoyo a sus ideas. Sin embargo, en su artículo de 1967 [12] y posteriormente en su carrera, Margulis se equivocó al descartar la evolución mediante la acumulación gradual de mutaciones beneficiosas y, en cambio, promover la endosimbiosis como principal fuerza impulsora de la evolución [7]. Entre sus posturas extremas sobre la endosimbiosis y la perspectiva gradualista de los neodarwinistas existe un punto intermedio razonable: que ambos modos de cambio evolutivo son compatibles con la evolución por selección natural.
¿Qué más se ha aprendido sobre el origen endosimbiótico de las mitocondrias y los cloroplastos en los casi 60 años transcurridos desde la propuesta de Margulis? Abundan nuevas teorías, siendo la más destacada la que sostiene que las mitocondrias se originaron a partir de una endosimbiosis entre los ancestros de una arquea del superfilo Asgard y una bacteria de un linaje conocido como alfa-proteobacteria (Figura 3). En el mundo actual, las alfaproteobacterias son muy comunes en todas partes, mientras que las arqueas de Asgard son conocidas por su capacidad de sobrevivir en ambientes extremos como fuentes hidrotermales.
La hipótesis general es que la endosimbiosis que dio lugar a las mitocondrias comenzó como una simbiosis metabólica, o sintrofia, entre las bacterias y las arqueas [15]. Sin embargo, la naturaleza exacta de la sintrofia que ocurrió entre estas bacterias y arqueas es incierta. La hipótesis del hidrógeno postula que la bacteria producía hidrógeno y dióxido de carbono como productos de desecho de su metabolismo, y que las arqueas consumían estos compuestos para sintetizar moléculas orgánicas [9] (Figura 4). Análisis filogenómicos más recientes de diferentes miembros de las arqueas Asgard sugieren que las arqueas metabolizaban nutrientes orgánicos para producir hidrógeno o compuestos ricos en hidrógeno como desechos, que luego eran utilizados por las bacterias como fuente de energía (Figura 4) [14]. En ambos casos, la sintrofia beneficiaba a la productora de desechos ricos en hidrógeno al evitar su acumulación y contaminación del ambiente, mientras que el consumidor obtenía energía "gratis".
La endosimbiosis que transformó a una alfa-proteobacteria ancestral en mitocondria fue un paso esencial en la evolución de las células eucariotas. En el proceso de convertirse en mitocondrias, el endosimbionte transfirió la mayor parte de su genoma al núcleo de la célula hospedadora, lo que definió su estatus como organelo en lugar de una bacteria de vida libre [5]. Las secuencias de estos genes son muy similares a las de las alfa-proteobacterias, lo que proporciona más evidencia del origen endosimbiótico de las mitocondrias. Esta es una evidencia más de que el ancestro celular de los eucariotes, que albergó al endosimbionte que se convirtió en mitocondria, surgió dentro del dominio arqueano de la vida, de la misma manera que las aves surgieron dentro del clado de los dinosaurios [6].
Más allá de la sintrofia inicial, persisten muchas preguntas sobre el origen endosimbiótico de las mitocondrias. Por ejemplo, ¿el hospedador arqueano se parecía a un Asgard moderno, o era más avanzado? ¿Podría haber tenido un núcleo y un sistema de endomembranas? La idea de que la endosimbiosis mitocondrial ocurrió antes de la formación del núcleo y las endomembranas se conoce como el escenario temprano de las mitocondrias. Tanto la hipótesis del hidrógeno como el modelo de flujo inverso apoyan este escenario temprano. Una visión opuesta es que la endosimbiosis ocurrió después de que la célula hospedadora ya había desarrollado uno o ambos organelos, una hipótesis conocida como escenarios tardíos de las mitocondrias.
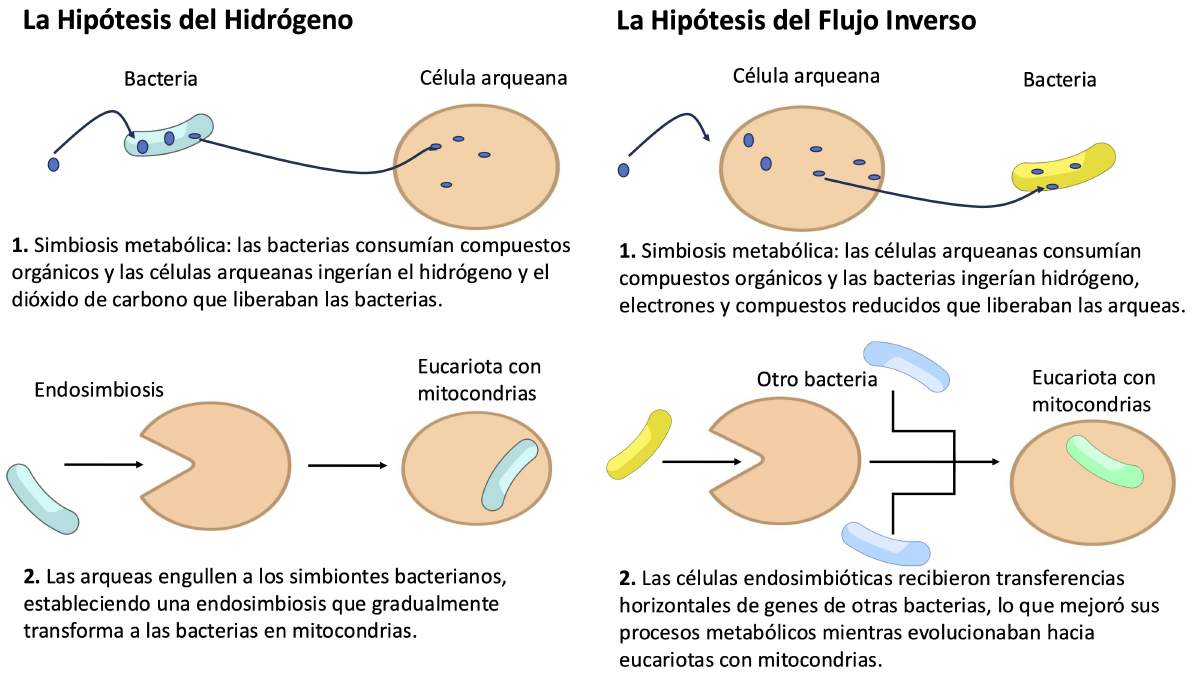
Figura 4. Dos hipótesis (de unas 30 que se han propuesto [10]) para explicar el origen de las mitocondrias como resultado de simbiosis metabólicas entre bacterias y arqueas. Modificado de [15].
¿Qué teoría es correcta? La respuesta depende en gran medida de si las mitocondrias eran esenciales para que las células generaran suficiente energía para desarrollar sus grandes tamaños, genomas complejos y otros organelos, así como para moverse y engullir procariotes como alimento o endosimbiontes [5]. Inicialmente, los defensores del escenario mitocondrial tardío propusieron que las mitocondrias eran esenciales para proporcionar la energía necesaria para la fagocitosis de un endosimbionte. Sin embargo, hoy sabemos de endosimbiontes bacterianos que viven dentro de otras bacterias, lo que sugiere que algún mecanismo de "engullir" endosimbiontes debe existir sin necesidad de fagocitosis (un proceso que las bacterias no pueden realizar) [4,5,13].
Por otro lado, los defensores del escenario temprano señalan que algunos eucariotes unicelulares actuales no dependen de metabolismos basados en fosforilación oxidativa —el tipo de metabolismo mitocondrial— o incluso han perdido las mitocondrias por completo, y parecen vivir perfectamente. Además, algunos de ellos pueden realizar fagocitosis. Aunque las arqueas Asgard comparten ciertas características con los eucariontes, no parecen poseer la capacidad de fagocitar. Esto sugiere que la evolución de la fagocitosis ocurrió después de la divergencia entre los eucariotes y sus ancestros arqueales, y que las mitocondrias no eran necesarias para la captura de endosimbiontes. Cómo un hospedador Asgard pudo engullir a un endosimbionte bacteriano sigue siendo una incógnita.
Tanto las mitocondrias como los cloroplastos se han integrado completamente en las células que los alojan. Aunque estos organelos se reproducen por fisión binaria, como las bacterias, solo pueden hacerlo dentro de las células que los albergan. Durante su transición de endosimbiontes a organelos, transfirieron la mayoría de sus genes al núcleo de la célula hospedadora, reteniendo solo un pequeño conjunto de genes esenciales para sus funciones metabólicas. Las mitocondrias conservan genes relacionados con la fosforilación oxidativa, y estos genes son similares a sus contrapartes en las alfa-proteobacterias. De manera similar, los genes de los cloroplastos codifican productos necesarios para la fotosíntesis, y el origen de estos genes se remonta a las cianobacterias.
Un ejemplo de cómo los endosimbiontes pueden pasar por la transición para convertirse en organelos lo ofrecen los nitroplastos de ciertas algas marinas. Estas estructuras fijadoras de nitrógeno fueron, en su origen, bacterias marinas fijadoras de nitrógeno que, hace aproximadamente 100 millones de años, comenzaron a residir dentro de las algas y parecen estar aún en proceso de transición de endosimbiontes a organelos [11].
Curiosamente, los genomas de las mitocondrias y cloroplastos contienen genes que provienen de bacterias distintas de las alfa-proteobacterias y cianobacterias. Estas bacterias pueden haber participado en eventos endosimbióticos con las células eucariotas en evolución, pero no se establecieron como organelos permanentes [1,15].
El advenimiento de la vida multicelular
La historia de la evolución de la vida es una de caminos ramificados, fusiones y experimentación. Una simbiosis entre procariotes que generó mitocondrias dio origen a las células eucariotas. Sin embargo, hubo un retraso de al menos 300 a 500 millones de años entre el surgimiento de las células eucariotas y la aparición de formas multicelulares simples, mientras que las formas complejas no surgieron sino hasta aproximadamente 1 a 1.2 mil millones de años después.
La multicelularidad fue una innovación evolutiva que proporcionó a las células eucariotas ventajas como la especialización, un mayor tamaño, una mayor complejidad, una movilidad mejorada y la capacidad de dominar muchos de los ecosistemas de la Tierra. Esta adaptabilidad contribuyó al auge evolutivo conocido como la explosión del Cámbrico, hace aproximadamente 540 millones de años. Durante este período, apareció una notable variedad de nuevos organismos que establecieron las bases para muchas de las líneas animales modernas [3].
Estas adaptaciones, que en última instancia surgieron de la endosimbiosis que permitió el desarrollo de la complejidad celular, no solo llevaron al surgimiento de estructuras como las columnas vertebrales, sino que también sentaron las bases para la asombrosa diversidad de formas de vida que vemos hoy en día. Desde los primeros organismos multicelulares hasta los complejos ecosistemas terrestres, la endosimbiosis marcó un punto de inflexión que transformó la historia evolutiva del planeta, demostrando cómo la cooperación puede ser tan poderosa como la competencia en la carrera por la supervivencia.
Referencias
- Archibald JM. 2015. Endosymbiosis and eukaryotic cell evolution. Current Biology. 25:R911-R921. http://dx.doi.org/10.1016/j.cub.2015.07.055.
- Dunn, M. F. 2025. Cómo la Vida se Volvió Compleja. Parte I: Los dominios de la vida y un poco de su historia evolutiva. La Unión de Morelos, 26 de mayo de 2025. https://acmor.org/publicaciones/c-mo-la-vida-se-volvi-compleja-parte-i-los-dominios-de-la-vida-y-un-poco-de-su-historia-evolutiva
- Gould, S. G. 1989. Wonderful Life: The Burgess Shale and the Nature of History. W. W. Norton and Co., New York, 347 p.
- Heidt A. 2022. The long and winding road to eukaryotic cells. The Scientist. 17 October 2022. https://www.the-scientist.com/the-long-and-winding-road-to-eukaryotic-cells-70556
- Lane N. 2015. The Vital Question: Energy, Evolution, and the Origins of Complex Life. W. W. Norton y Co., New York. 360 p.
- Maderspacher F. 2020. The long dark teatime of the cell. Current Biology 30,R451-R453 DOI: 10.1016/j.cub.2020.04.048
- Margulis, L. 2010. Symbiogenesis. A new principle of evolution rediscovery of Boris Mikhaylovich Kozo-Polyansky (1890–1957). Paleontol. J. 44, 1525–1539. https://doi.org/10.1134/S0031030110120087
- Marshall M. 2009. Timeline: The evolution of life. New Scientist. 14 July 2009; updated 27 April 2023. https://www.newscientist.com/article/dn17453-timeline-the-evolution-of-life/
- Martin, W and Müller, M. 1998. The hydrogen hypothesis for the first eukaryote. Nature 392:37-41. https://doi.org/10.1038/32096
- Raval, P. K., Zimorski, V., and Martin, W. F. 2004. Endosymbiotic theory. Encyclopedia of Evolutionary Biology Editor(s): Richard M. Kliman, , Academic Press, 2016, Pages 511-517,
- Romero D. 2024. Nitroplastos y diazoplastos, los nuevos organelos que surgieron de las aguas. La Unión de Morelos, 6 de mayo de 2024. https://www.launion.com.mx/blogs/ciencia/noticias/249465-nitroplastos-y-diazoplastos-los-nuevos-organelos-que-surgieron-de-las-aguas.html
- Sagan L. 1967. On the origin of mitosing cells. Journal of Theoretical Biology 14:225-274. https://doi.org/10.1016/0022-5193(67)90079-3
- Sousa, F. L., Neukirchen, S., Allen, J. F., Lane, N. & Martin, W. F. 2016.
Lokiarchaeon is hydrogen dependent. Nat. Microbiol. 4, 16034. DOI: 10.1038/nmicrobiol.2016.34
- Spang, A., Stairs, C.W., Dombrowski, N. et al. Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism. Nat Microbiol 4, 1138–1148 (2019). https://doi.org/10.1038/s41564-019-0406-9
- Wilcox C. 2019. Researchers rethink the ancestry of complex cells. Quanta Magazine April 9, 2019. https://www.quantamagazine.org/researchers-rethink-the-ancestry-of-complex-cells-20190409/
Esta columna se prepara y edita semana con semana, en conjunto con investigadores morelenses convencidos del valor del conocimiento científico para el desarrollo social y económico de Morelos.
